

记忆的本质是什么?
为什么可以不经你同意,任意删除记忆?

当你走在大街上看到一个漂亮的女孩,女孩的模样印刻在了你的脑海里。
一开始她是无比清晰的,但片刻之后,她开始变得模糊,紧接着只剩下一个轮廓,你越来越想不清她的样子……但也有可能她的面孔久久印刻在你的脑海里,甚至多年以后你还能记起那一瞥,清晰地好像刚刚发生。
不仅是那个女孩,你甚至还记得那片喧嚣的街区,温暖的阳光,铺满砖头的地面、空气里的味道。
……
为什么有时候,记忆会瞬息消失,但有时候却好像照片一样印刻在我们的脑海里,永远不会消失呢?

让我们一起来看看记忆究竟是什么
- 传统上,记忆被分成瞬时记忆、短时记忆、长期记忆。
1968年,心理学家Atkinson和Shiffrin提出经典的Atkinson-Shiffrin记忆模型(多重记忆模型),诠释了三种记忆的关系[1][2]。
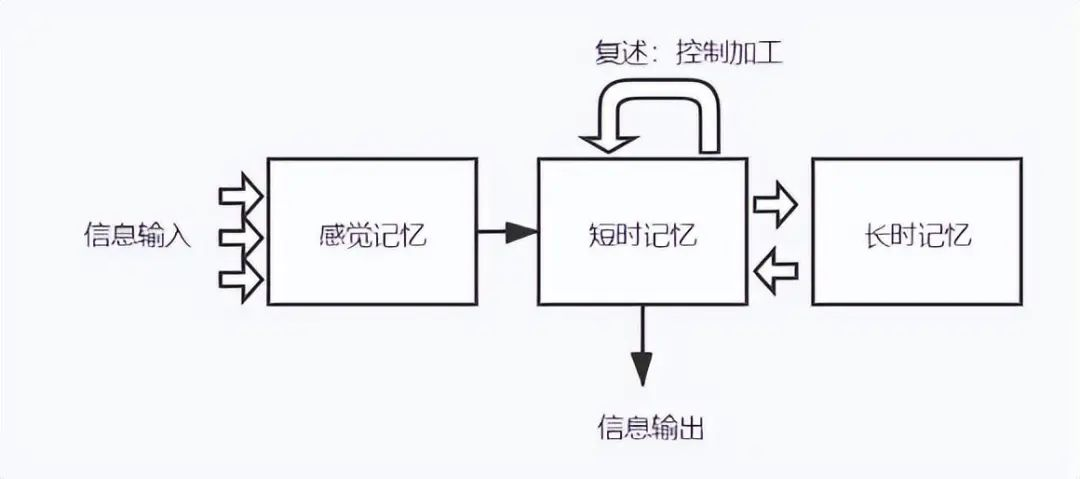
但由于三种记忆之间并没有明显的界限[3],所以也有研究者认为三种记忆没有明显的区分,属于单一模型,短时记忆只是长期记忆的临时激活[4]。
但就目前来说,多重记忆依旧是解释记忆的经典模型。
虽然该理论一开始并不完善,但经过几十年的发展,它已经形成了一个丰富的体系。
瞬时记忆(Sensory memory)
瞬时记忆也即感觉记忆,我们的感官在瞬间产生的记忆信息。
当我们走进一家餐厅,热闹的画面、嘈杂的声音、门口的凉风、猪蹄的味道……这些感觉通过神经末梢,传递到感觉中继站(丘脑为主),进行初步的整合、筛选,然后进入大脑皮层,形成感觉记忆。
感觉记忆是如何诞生的呢?
根据经典理论,视觉、听觉、味觉、嗅觉、触觉等五种感觉记忆的形成,都不尽相同。
视觉记忆
- 以下介绍的感官相关的记忆,皆是瞬时记忆。
视觉记忆持续的时间很短,大约只有几百毫秒。
我们的意识往往仅仅只能注意到瞬时记忆的一部分,例如当我们随意扫过天花板的时候,天花板上的普通纹路通常会被我们忽视(虽然这些瞬时记忆的确进入了大脑),仅仅在瞬间我们便忘了天花板上的纹路是什么形状。
然而,当天花板上的纹路和人类的面孔相似时,我们的脑海里就会留下深刻的印象。于是我们很清晰地记住了天花板上那个很像人类面孔的纹路,哪怕我们只看了一眼。

说穿了,瞬时记忆是没有认知参与的,当我们认知参与进去,对瞬时记忆进行再次加工时,它就自动变成了短期记忆。
视觉记忆究竟是如何形成的?
当我们看一眼一个陌生房间内场景,我们闭上眼睛,这个房间里的画面就会浮现在脑海里。心理学家乔治·斯佩林根据这个现象,设计出了一个实验[5],他让实验者站在一面镜子前辨识字母,而这些字母只会在很短的时间出现。
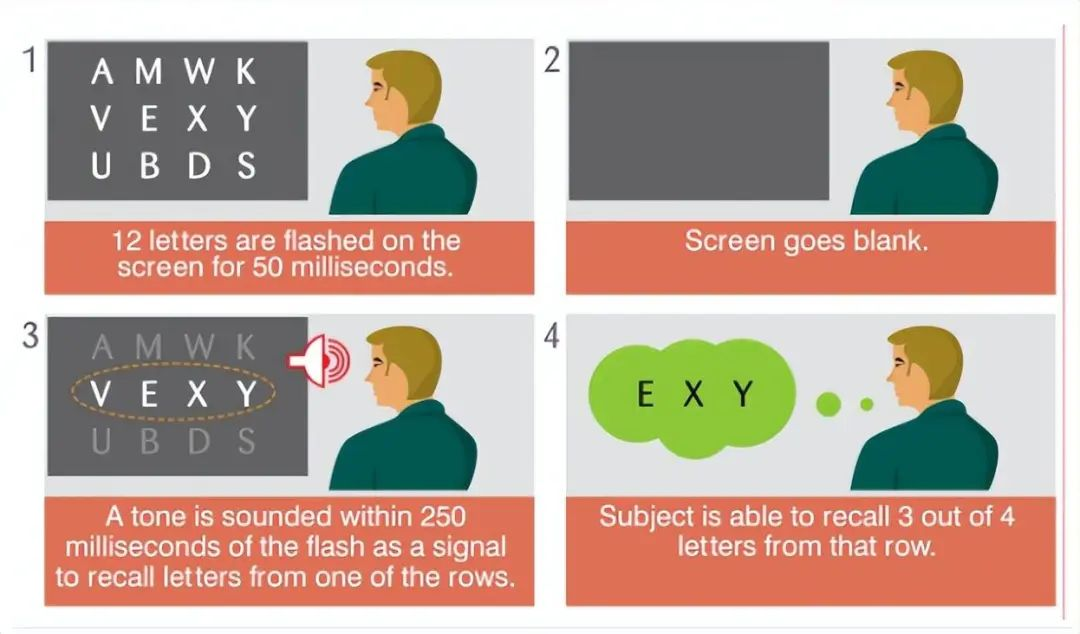
当出现的字母足够多的时候,参与者辨认字母的个数很快出现了上限,大多数人能辨识的字母个数不超过四五个。
乔治·斯佩林通过改进实验,把字母分行,发现参与者可以根据要求进行回忆。
这说明,视觉记忆的瞬间,所有的内容都是记住的,但在瞬间之后,大部分记忆都会被忘记,只会留下我们认知还能注意到的部分。当我们回忆这些记忆时,大脑中总是会出现当时看到的映象。
因此,乔治·斯佩林把这种瞬间形成的视觉记忆,称为映像记忆(iconic memory)[6]。

听觉记忆
听觉信息以声波的形式传播至耳朵里的毛细胞,然后转化成电信号,最终发送到颞叶听觉中枢进行处理。
听觉与视觉最大的区别在于,对于某个物体的视觉信号,我们的眼睛是可以反复扫描的。但某一个听觉信号,它可能出现后就不会再重复出现。而且听觉,往往是多个声音一起表达特定的信号或信息,必须听到下一个声音,上一个声音才可能变得有意义。
- 当你听一首歌的时候,回声记忆记录每个音符,然后大脑再连接起来,最终识别为一首歌。
所以听觉中枢处理记忆,与视觉中枢有所不同。
当我们听到一个声音后,这个声音似乎总是在脑海里回荡。
因此,在1967年认知心理学之父奈瑟尔(Ulric Neisser),提出了回声记忆(echoic memory)的概念。听觉记忆会在大脑内短期重复播放,并产生共鸣[7]。
回声储存的时间,不同的研究者有着不同的结果。通常在1~10秒范围内,一般认为4秒左右。不过也有研究表明,在没有其它信息竞争的情况下,回声记忆可长达20秒[8],这已经进入了短期记忆的范畴。

味觉记忆&嗅觉记忆&触觉记忆
无论味觉、嗅觉还是味觉记忆,与其它感觉记忆也是相似的。
几种瞬时记忆都受到不同中枢的控制,虽然因为不同的原理,各自有着不同的形成模式(例如触觉的振动同步激活[9])。但一开始都是形成高精度的信息,但会在数秒内迅速衰退。
感官记忆一经衰减或者被新的记忆取代,存储的信息就再也无法访问,并最终丢失。

除了基本的感官外,还有本体感觉记忆
手术麻醉后患者对肢体的位置会有“幻影”感知,同样被截肢的人也会有幻肢感,这种身体简化模型的感知,被成为“身体图式(body schema)”[10]。
研究表明,让受试者把手臂恢复到原来所在的位置,高精度记忆同样在2 ~4 秒快速衰减[11]。
这说明本体感觉,同样存在瞬时记忆。
短时记忆(Short-term memory)
在打电话的过程中,对方让你记住一个电话号码,你记下但并没有写在本子上。等到你挂掉了电话,却发现已经忘记了刚才的电话号码。
你刚刚明确记住电话号码时产生的记忆,便是短时记忆。
虽然绝大部分瞬时记忆都会在很短的时间消失,但被我们所注意到的信息,却可以被储存为短期记忆。
短期记忆被认为保留的时间为5-20秒,一般不超过1分钟。
早在1956年,心理学家乔治·米勒(George A. Miller)便进行实验发现,人类短期记忆储存的信息是有上限的。米勒给出的结果是7±2,称为米勒法则[12]。7这个数字,一时间也被带上了神秘色彩。但后来的研究表明,短期记忆的储存上限是4±1。
有一个与短时记忆相似的概念,叫做工作记忆(working memory)。

虽然在不少文章中,会把短时记忆等同于工作记忆,但二者其实具有很大的区别。
工作记忆指的是,对信息进行暂时加工和贮存的记忆系统。
如果把短时记忆比喻成输入电脑的临时数据,那么工作记忆就是电脑处理这些临时数据时,进一步产生的临时数据总和。
所以,工作记忆包含短时记忆,但短时记忆不能等同于工作记忆[13]。但在一些语境中,二者依旧存在混用的情况。尤其是进入21世纪以来,形成了研究工作记忆的热潮后,有时候短期记忆的概念也往往被工作记忆所代指。
工作记忆模型,由巴德利(Baddeley)和希奇(Hitch)在1974年提出,并在21世纪初得到完善[14]。
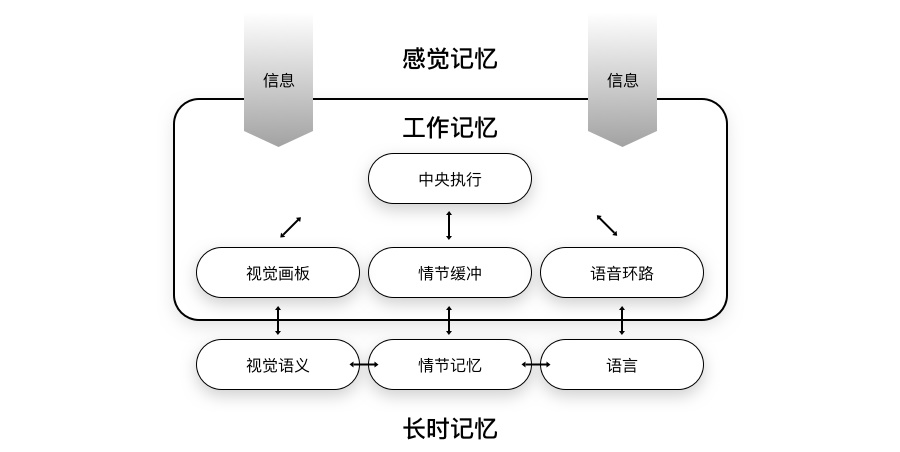
当你通过听、说、读、写尝试记住一串单词时,综合的感官信息会进入我们的大脑。当我们注意到要背的单词时,大脑会提取相应的关键信息,从而产生相应单词的短期记忆。
在这个过程中,「视觉画板」会加工视觉和空间信息,而「语音环路」则会记住单词的顺序。「中央执行」则在这个过程中,分配我们的注意力。「情节缓冲」则保持加工后的信息,支持后续加工。
那短期记忆产生的分子机制是怎么样的呢?
1949年,唐纳德·赫布(Donald Olding Hebb)提出著名的赫布理论[15]:
- 突触前神经元对突触后神经元进行反复且持续的刺激,便可以增强突触之间的链接。

该理论描述了突触可塑性的原理,反应了突触的学习和记忆机制。
当我们对单词进行学习和记忆,相关的突触连接就会增强,而当相关的知识被遗忘时,相关的连接就会减弱。
埃里克·坎德尔(Eric R. Kandel)通过海兔记忆的分子机制,证明短期记忆与长期记忆都发生在突触部位,并在2000年获得诺贝尔生理学或医学奖[16]。
突触可塑性分成短期可塑性和长期可塑性。
短期可塑性影响短时记忆,而长期可塑性影响长期记忆。
短期记忆形成的关键一环是蛋白质磷酸化。
而这个过程,是一个复杂的生物化学过程。
首先神经递质会促进神经元胞体制造环磷酸腺苷(cAMP),然后cAMP活化蛋白质活化酶A(PKA),PKA再使相应的离子通道蛋白质磷酸化,从而造成相应离子通道的失活。
- 例如,钾离子管道蛋白质磷酸化后,就会造成钾离子通道失活。
相应离子通道失活后,就会增加钙离子(Ca2+)的释放。现在的大量研究表明,这些钙离子来源于胞体内质网,它同时决定着钙离子的数量和分部[17]。
钙离子(Ca2+)是突触可塑性的核心介质。
它的大量释放会增加突触间神经递质的释放,从而短期增强突触连接,形成短时记忆。
长期记忆(Long-term memory)
当我们能记住一个单词几分钟,便进入了长期记忆的范畴。但有些长期记忆,我们只能记几个小时,有些却能记好几年,甚至一辈子……
这些过程,大脑中又发生了了什么呢?
虽然短时记忆有着容量上限,但长期记忆却是无限的。根据工作记忆理论,信息需要先进入工作记忆,才能储存到长时记忆中。长期记忆的储存速度,取决于工作记忆所能容纳的信息量。工作记忆的容量越大,学习速度越快[18]。
长期记忆的形成,需要经历两个关键步骤:
突触巩固(Synaptic consolidation)和系统整合(Systems consolidation)。
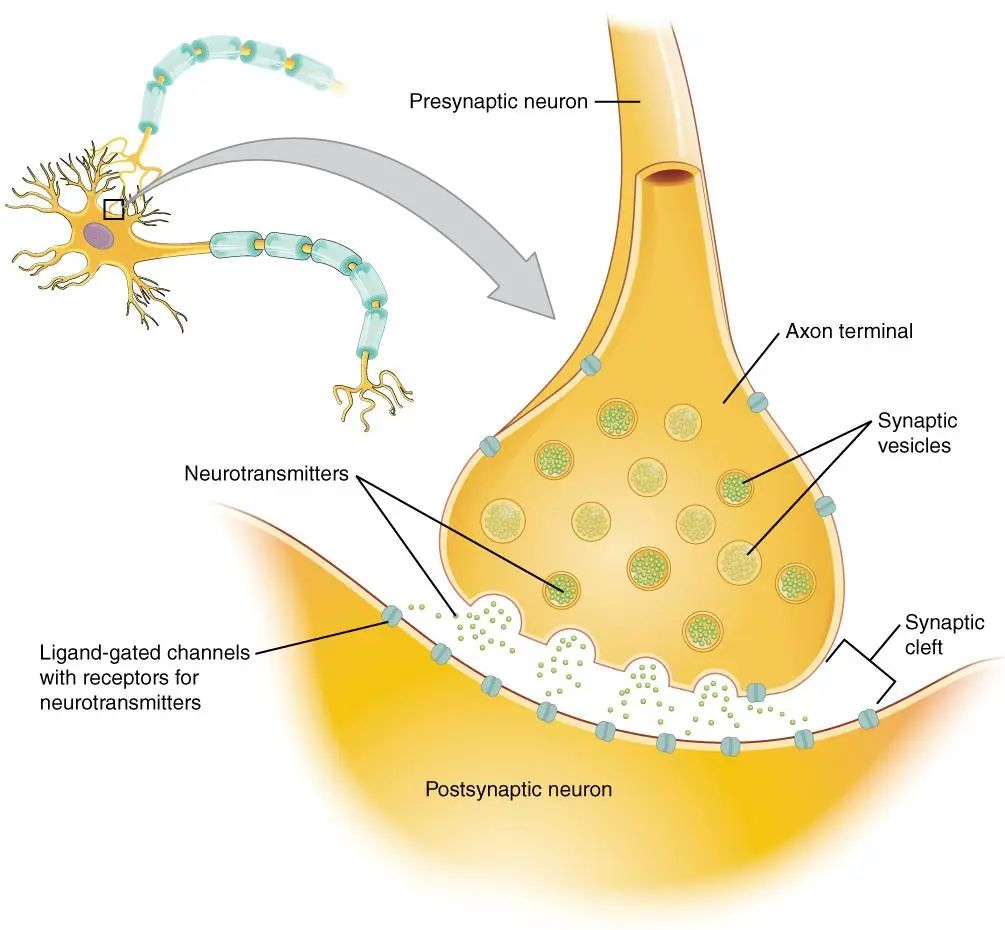
突触巩固(Synaptic consolidation)
突触巩固,主要依赖于突触的长时程增强(Long-term potentiation,LTP)[19],它也几乎是所有动物都具有神经机制,通常在几分钟、几个小时,但可以持续24小时以上[20]。
当然,相对于需要几周、几个月甚至几年的系统整合[21],突触巩固却是快了很多。
根据坎德尔(Kandel)的研究,突触的长时程增强(LTP)同样会发生蛋白质磷酸化,而且同样会经历“神经递质→cAMP→PKA→蛋白质磷酸化”的过程,从而增强突触连接。
但不同的是,这个过程之后神经元细胞核还会合成全新的式蛋白激酶——PKMζ[22]。
蛋白质的合成,是通过分子级联引起转录因子对DNA的甲基化而达成的。
研究表明,维持PKMζ的活性便能增强长期记忆,抑制PKMζ便能擦除已经建立的长期记忆,再消除抑制后,突触编码和存储新的长期记忆的能力还能再次恢复。
以上整个过程,并不会影响短期记忆。
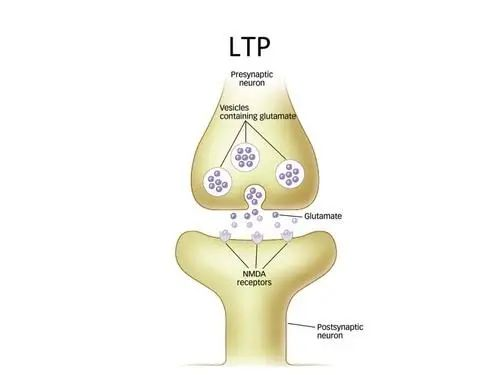
我们的记忆符合遗忘曲线,最可能的原因便是长期记忆依赖于蛋白质的合成。通过一定时间间隔的反复巩固(复习),就能让长期记忆越来越稳定[23]。
- 除此之外,脑源性神经营养因子(BDNF)对长期记忆的持久性也很重要。
虽然记忆的形成涉及到单个突触连接的增强,但记忆的储存方式却是整体编码的。
传统观点认为,记忆在形成过程中会产生记忆痕迹(memory trac)。
但实际,即便记忆痕迹存在,它也不是存在于单个突触连接中,而是由神经网络决定。
突触具有集结学习(Volume learning)机制,记忆和突触并非一一对应关系。
几十年前的研究就表明,通过少量切除猫脑不同组织,记忆不会受到影响。而切除的面积足够大之后,记忆才会受到影响。
可以认为,记忆痕迹是在神经网络中传播的,今天可能在这个突触上,明天可能就在另外一个突触上。一个大脑皮层神经元便拥有29800个突触,兴奋性突触后结构存在超过1000种蛋白,一个神经元的突触连接可能参与多个记忆痕迹。
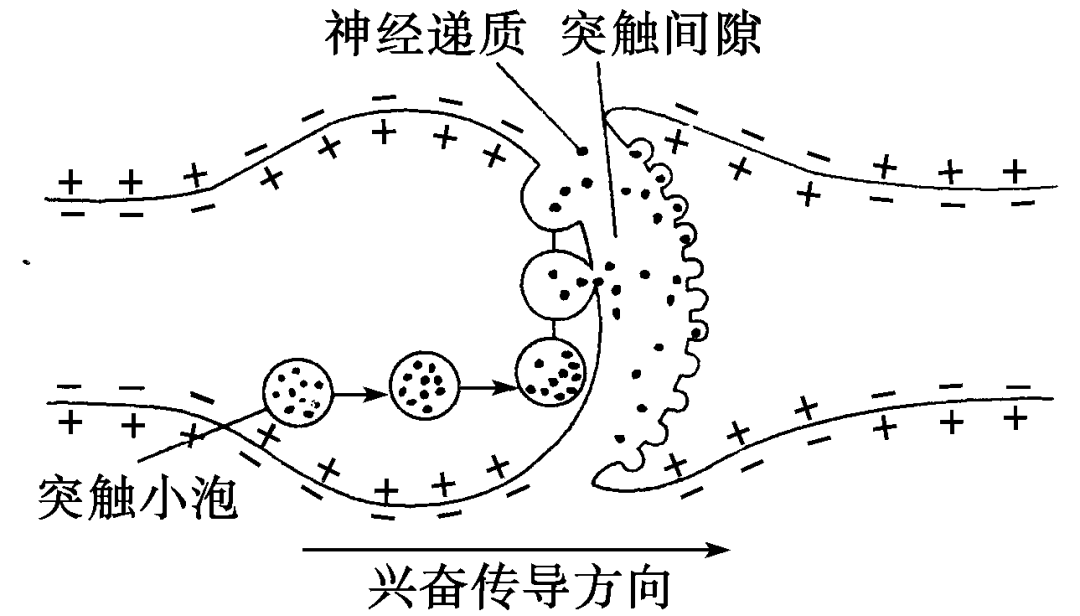
系统整合(Systems consolidation)
系统整合依赖于海马体的功能:以更持久的形式把记忆储存到新皮层内。

海马体成对存在于丘脑和内侧颞叶之间,3~4层脑皮层。
海马体
人类最开始对海马体记忆功能的认知,源于一场特殊的脑组织切除手术。
20世纪初,人类对大脑认知突飞猛进,在对待大脑疾病的治疗上,却有些野蛮而自负。这才有了莫尼兹在30年代发明前额叶切除手术,并得到广泛认同,且在1949年获得了诺贝尔奖。
虽然额叶切除术一直充满争议,但癫痫的手术治疗也是从那个时代发展起来的。
相对比起现在对病灶的精准切除,几十年前就有点随缘了。
1953年,为了治疗一位叫做莫莱森的27岁病人,医生切除了包括他海马体在内的部分边缘系统。虽然病人的癫痫得到了控制,但他的记忆却出现了问题。

H.M.病人
从1957年开始之后的几十年,他被当做典型来研究,称为“H.M.病人”。
他一直活到了2008年,对人类记忆的研究也算是做出了突出贡献[24]。
首先,术后的莫莱森出现严重的顺行性遗忘(无法形成长期记忆),以及部分逆行性遗忘(忘记手术之前的事情)。
他忘记了手术前短期内发生的任何事情,但记得多年前发生的事情,包括童年。
虽然短期记忆和工作记忆没有受到影响,但他无法形成情景记忆(包括时间和空间两个要素,例如我昨天8点在小区门口吃了豆腐脑这件事情,将永远不会再想起来)。
通过把药物注射进入大鼠的海马体,抑制突触增强时的蛋白质合成,研究者发现,虽然大鼠的学习能力没有表现出异常,但两天之后学习的内容就会完全遗忘。
在长期(永久)记忆形成过程中,随着海马向大脑皮层编码越来越多的信息,当回忆起这些信息时,它会加强大脑皮层与大脑皮层的联系,从而使记忆渐渐独立于海马体。
整个过程,涉及到海马体中多巴胺的释放和新蛋白质的合成[25]。
海马体也并非只有记忆功能,它还能编码来杏仁核的信息,而杏仁核主要负责人的情绪,这是我们能通过记忆通感、共情的根本原因[26]。
海马体中还存在“位置细胞(Place cells)”,可让海马体拥有“认知地图”的功能,可以让我们在回忆时想起去过的地点,以及如何去那个地方[27]。
总的来说,海马体相当于大脑中的记忆中继站,输入的各种感知构成记忆,然后在海马体中作为一个整体事件储存起来。最后再经过信息整合,输入大脑新皮层,从而形成永久记忆。
睡眠时的记忆整合,也是人类需要睡眠的重要原因之一[28]。
当我们回忆以前的记忆,记忆还会发生再巩固[29]。记忆再巩固的过程,还可能再次激活我们的各种感官皮层,产生身临其境的感觉,记忆也会经历一个保持、加强,再修改的过程。
不仅长期记忆可能变得更稳定,一些记忆偏差,或者无中生有的记忆塑造,也可能在这个过程发生。
虽然我们有时候很迷信自己的记忆,但其实,很多时候,记忆并没有那么可靠。

关于未来
总的来说,记忆更接近于一种“云”的方式储存在大脑中的,要实现数字化地存储记忆,需要同时弄清楚它的微观和宏观机制。虽然我们已经有了一定的认识基础,但对于完全弄清楚,依旧还比较遥远。
个人认为,记忆的数字化存储可能需要涉及到大量的容量和算力,需要超级计算和人工智能的互相结合才能实现。它会是一种对人类记忆的复刻。
相对来说,操控记忆会显得更容易一些,甚至现在便可能做到初级一些的操控记忆。例如可以充分利用记忆的可塑性,改变感官信息,让受试者不相信已经发生过的事情,或者对未发生过的事情深信不疑。
至于移植记忆,恐怕难度会是最高的。
如果记忆的数字化是一种超级电脑内的复刻,那么移植记忆涉及到的却是记忆的再输入、再创造。
对于一个空白的大脑来说,例如现在克隆一个完全一模一样的我,在大脑发育成长的过程中,把我生平的记忆,从逐渐输入他的大脑,建立和我基本一致的神经网络,相关记忆才算移植成功。
但如果要把我的记忆完全输入屏幕前的任何一个你的大脑中,就需要你大脑中有和我记忆相近的神经网络,然而每个人的大脑都是根据自己的生平记忆,进行强化和巩固的。
记忆的移植,可能也意味着需要找到什么方法,对大脑微观层面进行改造(例如,超级人工智能控制的纳米机器人,做到精准的突触修剪和巩固)。
同时,考虑到记忆移植对自我意识的影响,记忆移植也可能需要一个十分缓慢的过程。
- 当然,也有一个讨巧的方式,那就是利用影音设备详细记忆每个人生平的细节(要求不高时,这一步也可以省略),然后在储存记忆的过程,也同时储存相应的感官信息。当进行“记忆移植”的时候,进行可视化的输入,犹如元宇宙再体验的过程。当然,这其实也算不上真正意义上的记忆移植,只是“亲历”别人的记忆。但即便是这种技术,对于现今的人类都是遥不可及的。
所以,未来数字化地存储、操控和移植人类记忆并非不可能,但却十分遥远。
参考文献
[1] Atkinson R C , Shiffrin R M . HUMAN MEMORY: A PROPOSED SYSTEM AND ITS CONTROL PROCESSES 1[J]. Human Memory, 1977:7-113.
[2] Öğmen Haluk, Herzog M H . A New Conceptualization of Human Visual Sensory-Memory[J]. Frontiers in Psychology, 2016, 7:830.
[3] Brown G D A , Neath I , Chater N . A ratio model of scale-invariant memory and identification. 2007.
[4] Cowan, Nelson. [Progress in Brain Research] Essence of Memory Volume 169 || Chapter 20 What are the differences between long-term, short-term, and working memory?[J]. 2008:323-338.
[5] Sperling G . The Information Available in Brief Visual Presentation[J]. Psychological Monograph, 1960, 74.
[6] Sperling G . A Brief Overview of Computational Models of Spatial, Temporal, and Feature Visual Attention[M]. 2018.
[7] Radvansky G A . Human Memory: Second Edition[M]. 2015.
[8] Glucksberg S , Cowen G N . Memory for nonattended auditory material[J]. Cognitive Psychology, 1970, 1(2):149-156.
[9] Davis K D , Kwan C L , Crawley A P , et al. Functional MRI Study of Thalamic and Cortical Activations Evoked by Cutaneous Heat, Cold, and Tactile Stimuli[J]. Journal of Neurophysiology, 1998, 80(3):1533.
[10] Bromage P R , Melzack R . Phantom limbs and the body schema[J]. 1974, 21(3):267-274.
[11] Gentili M E , Verton C , Kinirons B , et al. Clinical perception of phantom limb sensation in patients with brachial plexus block[J]. European Journal of Anaesthesiology, 2002, 19(02):105-8.
[12] Miller G A . The magical number seven, plus or minus two : Some limits on our capacity for processing information[J]. Readings in Mathematical Psychology, 1963, 1.
[13] Jonides J , Lewis R L , Nee D E , et al. The Mind and Brain of Short-Term Memory – Annual Review of Psychology, 59(1):193[J]. Working Memory Attention Encoding Storage Retrieval, 2007.
[14] Baddeley A . The episodic buffer: a new component of working memory?[J]. 2000, 4(11):0-423.
[15] Hebb, D.O.The Organization of Behavior. New York: Wiley & Sons.1949.
[16] Eric, R, Kandel. NOBEL LECTURE: The Molecular Biology of Memory Storage: A Dialog Between Genes and Synapses[J]. Bioscience Reports, 2001.
[17] Justin K. O’Hare et al. Compartment-specific tuning of dendritic feature selectivity by intracellular Ca2+ release. Science, 2022.
[18] Nikoli D , Singer W . Creation of visual long-term memory[J]. 2007, 69(6):904-912.
[19] Bliss T V P . Long-lasting potentiation of synaptic transmission in the dentate area of the anaesthetized rabbit following stimulation of the perforant path[J]. The Journal of Physiology, 1973, 232.
[20] Dudai Y . the neurobiology of consolidations, or, how stable is the engram?[J]. 2017.
[21] Yadin, Dudai, Avi, et al. The Consolidation and Transformation of Memory[J]. Neuron, 2015.
[22] Costamattioli M , Sonenberg N . Translational control of gene expression: a molecular switch for memory storage.[J]. Elsevier Science & Technology, 2008.
[23] Litman L , Davachi L . Distributed learning enhances relational memory consolidation.[J]. Learning & Memory, 2008, 15(9):711-6.
[24] Pattanayak R D , Sagar R , Shah B . The study of patient henry Molaison and what it taught us over past 50 years: Contributions to neuroscience[J]. 2014.
[25] Initial memory consolidation and the synaptic tagging and capture hypothesis[J]. European Journal of Neuroscience, 2020.
[26] Gluck M A , Mercado E , Myers C E . Learning and Memory: From Brain to Behavior[J]. Worth Publishers, 2008.
[27] Manns J R , Eichenbaum H . A cognitive map for object memory in the hippocampus.[J]. Learning & Memory, 2009, 16(10).
[28] Wamsley, J Erin, Tucker,等. Dreaming of a Learning Task Is Associated with Enhanced Sleep-Dependent Memory Consolidation.
[29] Tronson N C , Taylor J R . Molecular mechanisms of memory reconsolidation[J]. Nature Reviews Neuroscience, 2007, 8(4):262.
文章来源:瞻云微信公众号